Факультативные анаэробы
Escherichia coli, Citrobacter freundii, Serratia marcescens, Entrobacter aerogenes, Photobacterium phosphoreum, Leuconostos dextranicum, Bacillus polymuxa, Bac. maceruns, Acvomonas hydrophila.
Выделять водород способны также многие другие строго или факультативно анаэробные микроорганизмы [163].
Микроорганизмы из субстратов различной химической природы способны выделять разное количество водорода. При этом образование водорода может идти из соединений углеводного характера, включая крахмал и целлюлозу, из амино- и кетокислот. В табл. 11 приведены количества водорода в молях, выделяющегося при конверсии одного моля субстрата, и вычисленные на основе этих данных коэффициенты преобразования энергии (использованы данные работ [163, 166, 167]).
![Таблица 11. Коэффициенты преобразования энергии некоторыми водородпродуцирующими микроорганизмами [162]](pic/000021.jpg)
Таблица 11. Коэффициенты преобразования энергии некоторыми водородпродуцирующими микроорганизмами [162]
КПД преобразования энергии рассчитывался по уравнению
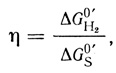 (1.3)
(1.3)где ΔGH20′ - свободная энергия окисления кислородом выделившегося водорода; ΔGS0′ - свободная энергия окисления кислородом субстрата. При вычислении свободных энергий окисления использовали термодинамические данные, приведенные в работе [164]
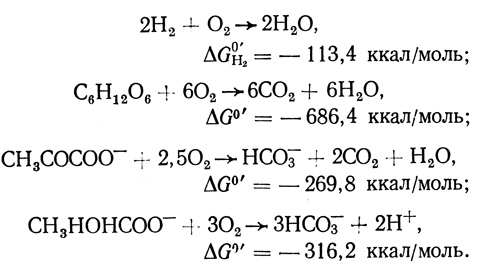 (1.4)
(1.4)Из таблицы 11 видно, что коэффициенты преобразования энергии в обычных условиях проведения процесса не превышают 20%.
Известно несколько метаболических путей образования водорода. Одна из возможностей, которая реализуется у факультативных анаэробов, например Е. coli, связана с разложением формиата под действием ферментативного комплекса формиатгидрогенлиазы [168]
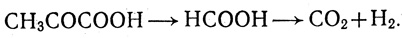 (1.5)
(1.5)Формиат обычно образуется из пировиноградной кислоты [165, 169]. Второй возможный путь связан с дегидрогенизацией НАДН или НАДФН [170-173]. Механизм такого рода обнаружен при исследовании бесклеточных препаратов C. kluyveri, также у ряда других бактерий
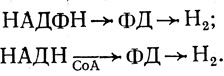 (1.6)
(1.6)Наиболее изучен метаболизм строгих анаэробов, выделение водорода у которых исследовано в работах [165, 169, 171, 176]. Конверсия глюкозы клетками некоторых клостридиев протекает в значительной степени с образованием уксусной и масляной кислот, СО2 и водорода [177, 178]. Материальный баланс в системе глюкоза-кислоты-водород-СO2 в первом приближении может быть представлен уравнением [167]
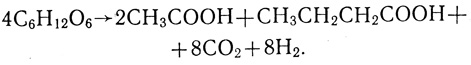 (1.7)
(1.7)Предполагается, что процессы образования масляной и уксусной кислот протекают параллельно и пути метаболизма имеют разветвления [164]. В целом стехиометрия процесса может быть представлена суммой двух реакций:
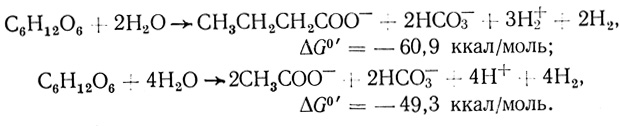
Схема метаболизма при образовании водорода в C. pasteurianum представлена на рис. 8 [164]. Выделение водорода в этом случае идет через восстановленную форму НАДН, которая образуется НАД - специфической глицеральдегидфосфатдегидрогеназой. Термодинамические аспекты образования водорода из НАДН в микробных системах рассмотрены в работах [178, 179].
![Рис. 8. Схема метаболизма при образовании водорода клетками Clostridium [164]](pic/000029.jpg)
Рис. 8. Схема метаболизма при образовании водорода клетками Clostridium [164]
На основе данных о метаболизме глюкозы при брожении, вызываемом клостридиями, представляет интерес вычислить коэффициенты преобразования энергии. Предельный КПД конверсии может быть получен, если расщепление глюкозы протекает по уравнению
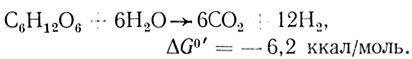
В этом случае 99% энергии конвертировано в водородную форму.
Но процесс протекает по уравнениям (1.7) и (1.8). Соответственно выход водорода существенно ниже. При образовании ацетата на моль глюкозы образуется 4 моля водорода. Это предельная величина, которая наблюдалась экспериментально [180, 181]. Образование 4 молей водорода соответствует 33% степени конверсии энергии. В случае процесса, направленного на образование бутирата (схема 1.8, 2 моля Н2), КПД преобразования равен 16,7%. Это определяется тем, что значительная энергия остается в алифатических кислотах. Таким образом, основная проблема при создании систем конверсии энергии биомассы в водород связана с превращением этих метаболитов в топливную форму. В этом плане работа может быть направлена как в сторону дальнейшего диспропорционирования и превращения их в водород, так и в сторону конверсии кислот в приемлемое углеводородное топливо.
Терминальными ферментами образования водорода у хемотрофных микроорганизмов являются гидрогеназы, которые известны давно. Однако в последние годы общенаучный и практический интерес к ферментам, осуществляющим образование водорода, заметно вырос. Этот интерес обусловлен исследованием физико-химического механизма действия ферментов, созданием гетерогенных катализаторов гидрогенизации - дегидрогенизации, в том числе для топливных элементов, особенностями действия этого класса ферментов в процессе образования или поглощения молекулярного водорода микроорганизмами. Молекулярные аспекты катализа гидрогеназами детально рассмотрены ниже.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://chemlib.ru/ 'Библиотека по химии'