Так что же такое ген?
Как только стало ясно, что в ядрах клеток сосредоточена наследственная информация и что эта информация дискретна и передается в виде неизменных порций - генов в различных их комбинациях, генетики широким фронтом развернули эксперименты, из которых стали вырисовываться количественные закономерности, управляющие наследственностью с ее изменениями. Было обнаружено, что действуя на хромосомы различными излучениями и химически активными веществами, можно вызвать мутации, т. е. изменения наследственных признаков. Все хромосомы выглядят под микроскопом не совсем одинаково. Сопоставляя вид хромосом с изменениями внешних признаков, удалось построить карты генов - генетики уже точно знали, где, на какой хромосоме, в каком месте располагается тот или иной ген, например ген окраски глаз или формы крыльев, у плодовой мушки - дрозофилы. Но молекулярная природа генов все еще не была известна, не было понятно, каким образом воспроизводится наследственный материал.
В 1927 г. русский биолог Н. К. Кольцов в статье "Наследственные молекулы" высказал предположение, что в ядрах клеток, а точнее - в хромосомах, находятся длинные полимерные молекулы. Вдоль такой молекулы и располагаются отдельные участки, управляющие передачей тех или иных признаков, - гены.
Кольцов предположил далее, что перед делением клетки каждая молекула, несущая гены, выстраивает рядом точную копию самой себе, после чего молекулы расходятся: оригинал в одну из клеток, копия - в другую. "Сложные молекулы не могут создаваться в организме заново", - писал Кольцов. И дальше: "Всякая, конечно, сложная органическая молекула возникает из окружающего раствора только при наличии уже готовой молекулы, причем соответствующие радикалы помещаются... на те пункты имеющейся налицо и служа- щей затравкой молекулы, где лежат такие же радикалы". Таким образом, Кольцов очень проницательно отметил, что наследственная информация на молекулярном уровне может передаваться подобно тому, как происходит кристаллизация веществ из раствора на затравке из кристаллов. Фактически в этом и состоит матричный принцип синтеза новых биополимерных молекул на пред существующих молекулах того же типа. К сожалению, в 20-е годы роль ДНК в передаче наследственности была еще совершенно неясна, и Кольцов считал, что вещество наследственности - это белок, содержащийся в хромосомах. Представление о том что гены - это молекулы особых белков, - просуществовало довольно долго, и окончательно от него отказались лишь Уотсон и Крик в начале 50-х годов.
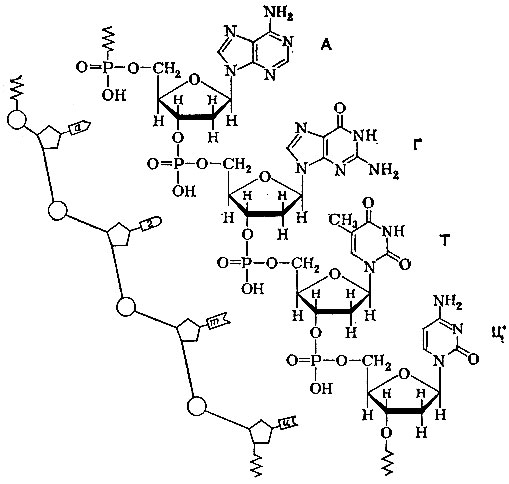
Здесь изображен кусочек ДНК, состоящий всего из четырех 'букв' - нуклеотидов. Слева - более простое условное изображение того же фрагмента. Кружками обозначены фосфаты, пятиугольниками - дезоксирибоза. Основания изображены различными фигурами, смысл которых станет ясен из дальнейшего. Маленькими буквами обозначены основания: а - аденин, г - гуанин, т - тимин, ц - цитозин, большие буквы - соответствующие им нуклеотиды: А, Г, Т, Ц
Что же было известно до Уотсона и Крика и что нового дала их работа? Прежде всего, они были твердо убеждены, что именно ДНК - носитель наследственной информации. Было известно, что молекула ДНК имеет очень большую молекулярную массу и что это, по-видимому, длинная цепь, образованная из множества нуклеотидов. Нуклеотиды соединяются между собой через фосфатные остатки - один фосфат при-соединен к двум гидроксильным группам, принадлежащим двум остаткам сахаров соседних нуклеозидов. Последовательность нуклеотидов - это первичная структура ДНК. Далее, было известно, что в ДНК со-держание аденина точно равно содержанию тимина, а количество гуанина точно равно количеству цитозина. Это так называемые правила Чаргаффа, установленные в конце 40-х -начале 50-х годов. Наконец, Крик располагал ренгенограммами образцов ДНК, полученными Розалиндой Фрэнклин.
Нужно было на основании этих данных решить, как же устроена молекула ДНК, как она расположена в пространстве и каким образом, почему, благодаря каким особенностям ДНК является веществом наследственности (а в это Уотсон и Крик верили свято).
С какой стороны подступиться, с чего начать? Очень импонировал обоим ученым путь, который привел Лайнуса Полинга к открытию знаменитой α-спирали белковой молекулы. Уотсон пытался понять, как именно Полинг открыл α-спираль. Вот что писал Уотсон.
Рентгенограма кристалла ДНК, получения Р. Фрэнклин. Расчет Крика показывает, что такую рентгенограмму может дать спиральная молекула
"Вскоре я усвоил, что Полинг достиг этого, опираясь больше на здравый смысл, чем на сложные математические выкладки. В своих рассуждениях он иногда оперировал уравнениями, но и тут в большинстве случаев можно было бы обойтись словами. Ключ к успеху Лайнуса надо было искать в том, что он доверился простым законам структурной химии, α-Спираль была открыта не с помощью простого созерцания рентгенограмм; главный фокус состоял в том, чтобы задать себе вопрос: какие атомы предпочитают соседствовать друг с другом? Основными рабочими инструментами были не бумага и карандаш, а набор молекулярных моделей, на первый взгляд напоминающих детские игрушки.
Мы не видели, что могло бы помешать нам решить проблему ДНК таким же способом. Нужно было только сконструировать набор молекулярных моделей и начать играть ими".
Вот так, "играя" с моделями, Уотсон и Крик и открыли свою знаменитую двойную спираль. Крик, как физик, произвел кое-какие расчеты и показал, что рентгенограмма кристаллической ДНК, полученная Фрэнклин, лучше всего соответствует именно спиральной укладке длинной молекулы в виде двух переплетающихся цепей.
Как же соединяются две нити ДНК, какие силы удерживают их в переплетенном состоянии? Силы те же, что и в структуре Полинга - водородные связи.
Уотсон, собирая модели нуклеотидов и так, и этак в разных сочетаниях, нашел, что идеальным вариантом является такой, когда фосфатно-сахарный остов образует основу каждой из нитей, а азотистые основания обращены внутрь. Причем против аденина должен располагаться только тимин, а гуанин прекрасно подходит к цитозину. Два основания каждой такой пары соединены водородными связями. Аденин и тимин двумя связями, а гуанин и цитозин - тремя. Сразу нашло объяснение правило Чаргаффа: в молекуле ДНК А = Т и Г = Ц.
По своим размерам аденин настолько хорошо подходит к тимину, а гуанин - к цитозину, что получающаяся двунитевая спираль имеет по всей длине совершенно правильную конфигурацию, и толщина спирали в любом месте одинакова (диаметр ее составляет 2 нм).
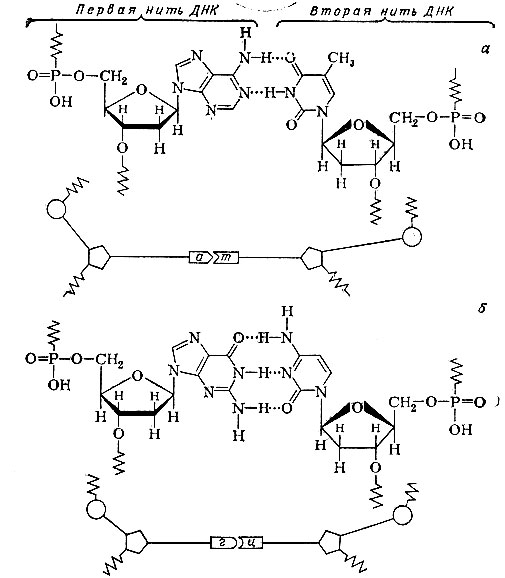
В молекуле ДНК против аделина одной цепи двойной спирали обязательно располагается тимин другой цепи (а). Такие же отношения существуют между гуанином и цитозин (б)
Теперь легко объяснить, как происходит воспроизведение молекул ДНК: молекула расплетается на две нити, на каждой нити строится вторая дополнительная к ней нить, и обе образовавшиеся двунитевые молекулы расходятся в две дочерние клетки.
Не нужно только думать, что две нити ДНК при репликации (т. е. при удвоении) расплетаются как веревка, составленная из двух прядей. С какой скоростью должны были бы вращаться в таком случае, например, концы нитей ДНК кишечной палочки, состоящей из трех миллионов пар нуклеотидов? На каждый виток приходится 10 пар азотистых оснований. Клетка кишечной палочки делится каждые 20 мин, т. е. за это время должна раскрутиться ДНК длиной 1 мм (именно такова длина трехмиллионно нуклеотидной ДНК кишечной палочки). Получается, что скорость расплетания должна была бы составлять 15 000 оборотов в минуту. Это совершенно немыслимая скорость. Приходится предположить, что при удвоении молекулы ДНК она сначала расщепляется во многих местах, так что наращивание второй молекулы идет одновременно по всей длине ДНК.
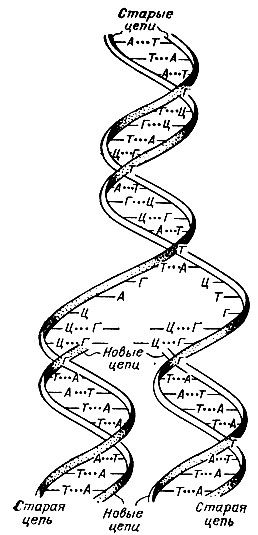
Так схематично можно представить процесс репликации молекул ДНК
Нам бы очень не хотелось, чтобы у читателя возникло впечатление, будто Крик и Уотсон пришли к своему открытию легко и просто. Были и мучительные поиски, и разочарования, и бессонные ночи. О том, как непросто все было, говорит хотя бы такой факт.
В разгар работы над структурой ДНК, когда многое еще было неясно, но кое-что в будущей структуре начало вырисовываться, Крику и Уотсону стало известно, что Лайнусу Полингу, творцу а-спиральной модели молекулы белка, кажется, удалось решить проблему структуры ДНК. Можно представить себе, как это известие взволновало Уотсона и Крика. Они принялись лихорадочно читать рукопись еще не опубликованной статьи и скоро поняли, что Полинг был далек от истины: его модель представляла собой трехнитевую спираль, в которой сахарофосфатный остов всех трех нитей находился в центре, а азотистые основания торчали в разные стороны. На этот раз?
Полинг, самый проницательный химик мира, как его охарактеризовал Уотсон, допустил ошибку.
Да, "просто, мудро, изящно", но вовсе не так легко, как это теперь нам кажется. Впрочем, на протяжении своей работы они всегда придерживались принципа - в природе все просто и изящно. Если им приходила в голову гипотеза, которая казалась недостаточно красивой и простой, они ее сразу отбрасывали.
Итак, был предложен красивый, простой, убедительный механизм "размножения" молекул ДНК. В каком соотношении он находится с геном? Сегодня мы знаем, что ген - это часть очень длинной молекулы ДНК.
В 1969 г. сотрудники Гарвардской медицинской школы в США под руководством Д. Беквита сумели выделить отдельный ген, "отстричь" его от остальной цепи ДНК. Этот ген находится в ДНК бактерии кишечной палочки и служит для синтеза фермента, который помогает бактерии усваивать сахар лактозу. Удалось даже рассмотреть выделенный ген под микроскопом. Его длина - 0,0015 мм, состоит он из 4700 пар нуклеотидов.
Вся же молекула ДНК, хранящая наследственные признаки сложных организмов, в сотни раз длиннее. Можно представить себе, какие сложные, какие длинные цепи молекул ДНК хранятся в ядре каждой из миллиардов наших клеток. Так, у млекопитающих в ядрах клеток содержатся ДНК, состоящие из 109 нуклеотидов. Ясно, что такими числами можно записывать огромное количество информации.
В то же время молекулы ДНК простейших вирусов относительно малы. Удалось взвесить и сфотографировать ДНК одного из бактериофагов. Эта ДНК представляет собой кольцо с длиной окружности около 2 мкм, состоящее из 5600 пар нуклеотидов. Молекулярная масса такой "крошечной" ДНК составляет 6 миллионов углеродных единиц. Функции бактериофага весьма примитивны, поэтому он и обходится та-кой короткой ДНК.
Как же реализуется информация, заложения в последовательности нуклеотидов в ДНК? Что, собственно, определяет эта последовательность? Оказалось, что последовательность нуклеотидов определяет только одно: последовательность аминокислот в Новосинтезируемых белках.
Фактически, текст, записанный четырех буквенным алфавитом из нуклеотидов, переводится в другой текст, записанный 20-буквенным алфавитом аминокислот. Каждый ген, т. е. определенный отрезок цепи ДНК, кодирует биосинтез одного белка (правило: один ген - один белок), точнее - одного полипептида, поскольку некоторые белки состоят из нескольких полипептидных цепей.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://chemlib.ru/ 'Библиотека по химии'